
Biological Control
We are currently studying the impacts of habitat complexity on arthropod communities, primarily natural enemies in cabbage fields, although arthropod communities in other vegetable crops (e.g. snap beans, squash, and sweet corn) are also being examined. Habitat complexity consists of living vegetational complexity and non-living structural complexity. We manipulate these two types of habitat complexity in the field between rows of vegetable crops and examine the impacts on community structure, arthropod abundance, and biological control using field based and molecular techniques. We suspect habitat complexity may provide refuge and additional resources for arthropods in agroecosystems. Manipulating habitat complexity may enhance the biological control services provided by the natural enemies naturally found in the field.
Pests in cabbage
Common pests in Michigan cabbage fields (see pictures below) include the imported cabbageworm (Pieris rapae L.), diamondback moth (Plutella xylostella L.), cabbage looper (Trichoplusia ni (Hübner)), cabbage aphid (Brevicoryne brassicae L.), striped flea beetle (Phyllotreta striolata (Goeze)), and onion thrips (Thrips tabaci (Lindeman)).

Natural enemies in vegetables
Diverse communities of natural enemies are found in Michigan vegetable fields, including generalist predators such as spiders, ground beetles, predatory stink bugs, lady beetles, minute pirate bugs, damsel bugs, predatory thrips, and big-eyed bugs. Specialized natural enemies, such as parasitic wasps and flies are also present. For instance, in cabbage parasitic wasps that attack the imported cabbageworm and diamondback moth: Cotesia rubecula and Diadegma insulare, are commonly found in the field.

Vegetational and structural complexity in cabbage
In 2010 and 2011, we examined arthropods in cabbage plots across different amounts of vegetational and structural complexity. Vegetational complexity was created in cabbage fields through manipulation of weed management intensity. Reduced weed management intensity led to greater weed pressures and greater vegetational complexity. Combining strip-tillage and a killed oat cover crop created mulch on the soil surface. Structural complexity was manipulated through oat cover crop kill date. Later kill dates resulted in greater mulch in the row-middle and greater structural complexity (see photos below).


Insect community structure
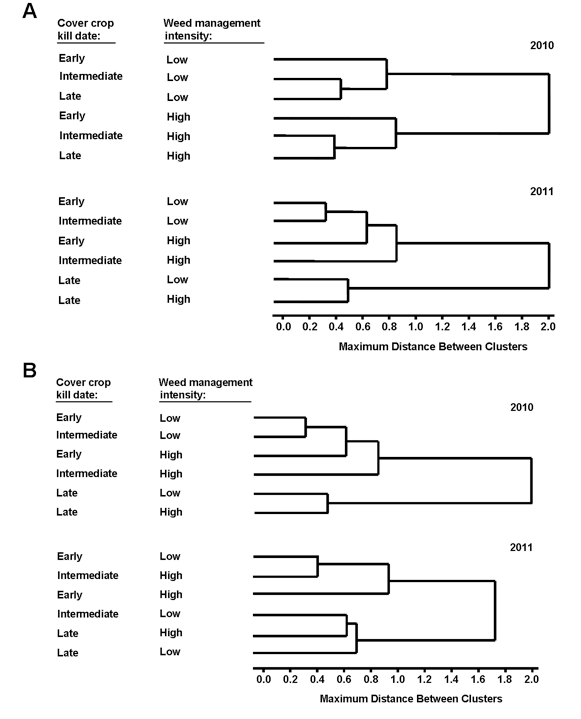
We performed cluster analysis to examine the impact of habitat structure on arthropod community structure. In 2010, herbivore communities were influenced by herbicide intensity (Fig. A). In 2011, when weed pressure was lower, herbivore communities were influenced primarily by oat kill date (Fig. A). Natural enemy communities were more consistently influenced by oat kill date than by herbicide intensity (Fig. B). It appears habitat complexity can shape both herbivore and natural enemy communities.
Insect abundance
In 2010 and 2011, we examined the impact of changes in vegetational and structural complexity on individual pest and natural enemy species. Greater vegetational complexity was associated with lower cabbage aphid abundance in both 2010 and 2011. Other pests were not consistently impacted by weed pressures. Several natural enemies, including predatory stink bugs, lady beetles, minute pirate bugs, and predatory thrips were positively impacted by vegetational complexity in at least one year.
Greater structural complexity led to greater imported cabbageworm and cabbage looper abundance in 2010. However, these trends were not observed in 2011. Natural enemies, primarily lady beetle species were positively impacted by structural complexity. One natural enemy species, Diadegma insulare, was negatively impacted.
Habitat complexity appears to positively impact arthropods in general, suggesting row-middle habitats may provide refuge and additional resources. Species negatively impacted by habitat complexity may suggest these row middle habitats cause interference of chemical or visual searching. These potential mechanisms are currently being explored by the lab.
Biological control
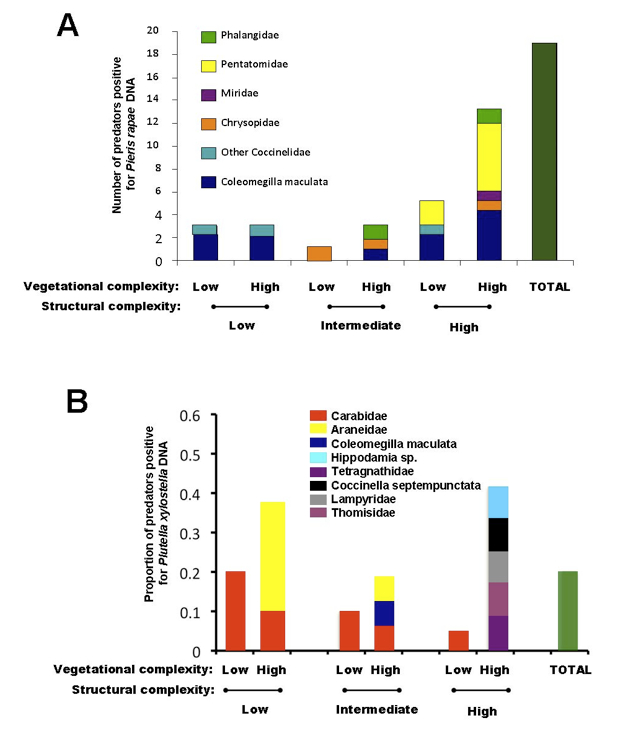
In 2010 and 2011, biological control was measured in our plots using molecular gut content analysis. This method detects prey DNA (e.g. Pieris rapae DNA and Plutella xylostella DNA) in the guts of field collected predators and shows predation rates within the field and identifies significant predators. In 2010, predation of the imported cabbageworm (Pieris rapae) was greatest in our plots with the greatest habitat complexity (Figure A below). The diversity of predators found positive for prey DNA in their guts was also greatest in these parts of the field. In 2011 (Figure B below), predation of another cabbage pest, the diamondback moth (Plutella xylsotella), was higher in plots with greater vegetational complexity. Structural complexity did not appear to influence predation of the diamondback moth in 2011. It appears that predation may be impacted by habitat complexity between cabbage rows. Possible mechanisms for this trend include that these habitats support a larger natural enemy community, enhance foraging efficiency of natural enemies, or reduce intraguild predation.
Manipulating structural complexity between cabbage rows
In 2012, we examined the impact of different forms of structural complexity on cabbage arthropods. Treatments included combinations of different mulch species (e.g. none, vetch, and rye) and the height of mulch (e.g. standing or crimped).


Insect community structure
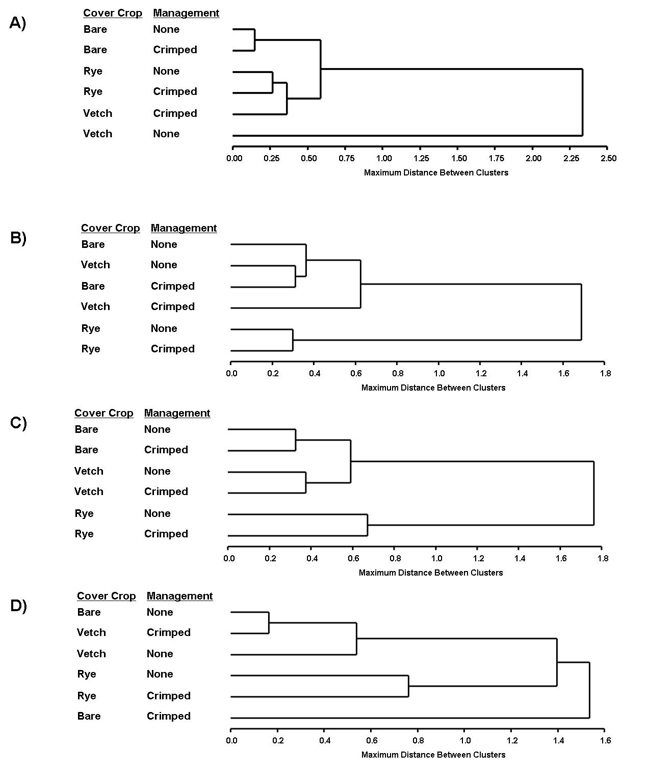
The herbivore community in 2012 was shaped by the cover crop species between the cabbage rows (Figure A). Natural enemy community dendograms show the predator community (Figure B), the ground predator community (Figure C), and the parasitoid community (Figure D) also being shaped by cover crop species, with natural enemy communities often distinct in plots with rye mulch. Crimping or the height of the mulch did not appear to shape our arthropod communities. In general, natural enemy communities were similar across vetch and bare soil plots. This is not surprising since rye and vetch plots were indistinguishable later in the season, due to the quick decomposition of vetch mulch. It appears the type of structural complexity provided by different mulch species (e.g. broadleaf vs. grass) influences community structure.
Arthropod abundance
The height of the cover crop mulch did not appear to influence arthropod abundance. However, arthropod abundance differed across cover crop species. Fewer flea beetles were observed in rye plots compared to vetch and bare soil plots. Other pest species were not different across treatments. Natural enemies, such as ground beetle species (e.g. Harpalus pennsylvanicus) and a specialized parasitoid, Cotesia rubecula, were also more abundant in rye plots. Our findings suggest rye mulch can enhance natural enemy abundance. Since rye decays more slowly, this mulch provides habitat and resources for a more extended period of time than vetch or bare soil plots.
Biological control
Using predator exclusion cages with sentinel prey (e.g. imported cabbageworm) indicates predation and parasitism rates within the field due to guilds of natural enemies (e.g. small predators and parasitoids and large predators). In 2012, manipulating structural complexity by changing mulch species or height did not influence predation (Figure A) or parasitism rates (Figure B) of the imported cabbageworm. However, we did observe 40-70% predation across our cabbage fields, with most predation due to small predators (lady beetle larvae, lacewing larvae, small spiders, etc.). Parasitism rates approached 25% during the later part of the cabbage growing season. This project indicates the importance of small natural enemies and the services they provide in our fields as natural biological control.

This work is part of A. Bryant’s graduate research. Article written by A. Bryant.